种群数量增长和数学曲线
种群的数量是指在一定面积或容积中某个种群的个体总数。一个种群的个体数目多少,也叫种群大小。理论上认为,种群大小决定于三个因素:(1)起始种群个体数量;(2)出生和死亡;(3)迁入和迁出。所有能影响种群的出生率、死亡率和迁移率的因素,都会影响种群数量的变化。种群数量的类型有以下几种。
一、种群在无限环境中的指数增长
这样的种群增长模型可以概括为两种:世代不相重叠种群的离散增长模型和世代重叠种群的连续增长模型。
种群在“无限”的环境中,即假定环境中空间、食物等资源是无限的,且气候适宜、没有天敌等理想条件下,种群的增长率不随种群本身的密度而变化,种群数量增长通常呈指数增长。
1. 世代不相重叠种群的离散增长模型
假定某种动物一年只生殖一次,寿命只有一年,那么,这种动物的种群就是世代不相重叠的。例如,草原上往往有季节性的小水坑,栖居在这些小水坑中的水生昆虫,雌虫每年产一次卵,卵孵化长成幼虫,蛹在泥中度过干旱季节,到第二年,蛹才变成虫,并交配、产卵。因此,这样的昆虫种群世代是不重叠的,种群增长是不连续的。对世代不相重叠的种群来说,假定种群增长不受资源、空间等条件的制约,没有迁入和迁出,没有年龄结构,那么,种群的增长就可以用下列方程表示:
![]()
其中N0为初始种群数量,t为时间,![]() 为种群的年增长率。
为种群的年增长率。
2. 世代重叠种群的连续增长模型
如果种群是世代重叠的,种群的增长则为不间断的连续增长。假定种群的其他特征与上述世代不相重叠的种群相同,种群的瞬时增长率为r,则种群仍表现为指数增长。
大多数种群的繁殖都要延续一段时间并且有世代重叠,即在任何时候,种群中都存在不同年龄的个体。这种情况要以一个连续型种群模型来描述,涉及微分方程。
其积分式为![]()
其中![]() 的定义与世代不相重叠种群增长模型的相同,e=2.71828……,r是种群的相瞬时增长率。
的定义与世代不相重叠种群增长模型的相同,e=2.71828……,r是种群的相瞬时增长率。
这个公式就是我们所需要的描述连续性变动种群动态的基本模型。这里r代表了种群的瞬时增长率(与密度无关)。如果r>0,种群上升;r=0,种群稳定;r<0,种群下降。用数学曲线表示如下图所示。
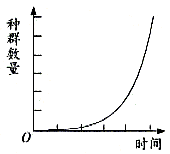
r值能表示物种的潜在增殖能力。例如恒温箱中培养细菌,从一个细菌开始,通过分裂,按2、4、8、16……增长,在短期中能表示出指数增长。许多具简单生活史的动物在实验培养中也具有类似的指数增长。在自然界中,一些一年生昆虫,甚至某些小啮齿类,在春季环境条件较好的情况下,其数量也会呈指数增长。自16世纪以来,世界人口表现为指数增长,所以一些学者称之为人口爆炸。
二、种群在有限环境中的逻辑斯谛增长
种群在有限环境中的增长,同样可以分为世代不相重叠种群增长和连续增长两类。种群在有限环境下的连续增长的一种最简单的形式是逻辑斯谛增长。
因为环境是有限的,大多数种群的指数增长都是暂时的,一般仅发生在早期阶段,种群密度很低,资源丰富的情况下,而随着密度增大,资源缺乏、代谢产物积累等,种群的增长率r势必降低。逻辑斯谛增长模型是建立在以下两个假设基础上的:
①有一个环境容纳量(通常以K表示),当![]() 时,种群为零增长,即
时,种群为零增长,即![]() ;
;
②增长率随密度上升而降低的变化是按比例的。最简单的是每增加一个个体,就产生1/K的抑制影响。例如K=100,每增加一个个体,就产生0.01影响,或者说,每一个体利用了1/K的“空间”,N个个体利用了N/K的“空间”,而可供种群继续增长的“剩余空间”只有(1-N/K)。
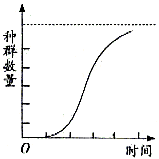
按此两点假设,密度制约导致r随着密度增加而降低,这与r保持不变的非密度制约型的情况相反,种群增长将不再是“J”型,而是“S”型(如下图所示)。“S”型曲线有两个特殊:①曲线渐近于K值,即平衡密度;②曲线上升是平滑的。产生“S”型曲线的最简单数学模型是在指数增长方程(![]() )上增加一个密度制约因子(
)上增加一个密度制约因子(![]() ),就得到生态学上著名的逻辑斯谛方程。
),就得到生态学上著名的逻辑斯谛方程。
![]()
在种群增长早期,逻辑斯谛增长与指数增长相差不多,种群数目N很小,N/K也很小,因此(1-N/K)接近于1,所以密度抑制效应可以忽略不计。但随着种群密度增大,密度抑制效应增加,直到当N=K时,种群就停止增长,种群达到了一个稳定的大小不变的平衡状态。
逻辑斯谛曲线常划分为五个时期:①开始期,也可称潜伏期,种群个体数很少,密度增长缓慢;②加速期,随着个体数增加,密度增长逐渐加快;③转折期,当个体数达到饱和密度一半(即K/2)时,密度增长最快;④减速期,个体数超过K/2以后,密度增长逐渐变慢;⑤饱和期,种群个体数达到K值而饱和。
逻辑斯谛模型的两个参数r和K,均具有重要的生物学意义。R表示物种的潜在增殖能力;K是环境容纳量,即物种在特定环境中的平衡密度。但应注意,K同其他生态学特征一样,也是随环境(资源量)的改变而改变的。
逻辑斯谛增长模型的重要意义:①它是许多两个相互作用种群增长模型的基础;②它也是渔业、林业、农业等实践领域中,确定最大持续产量的主要模型;③模型中两个参数r、K,已成为生物进化理论中的重要概念。